Нічне нетримання сечі, лікування енурезу у дитини
Нічне або денне нетримання сечі – поширена, неприємна та дуже травмуюча проблема. Психіка дитини може...
Різні видирецепторів шкіри (схема) 1 - вільні нервові закінчення з рогівки ока; 2 - дотичні пластинки Меркеля; 3 - дотичні тільця Мейсснера; 4 - нервове сплетення волосяної цибулини; 5 - кінцева колба Краузе; 6 - тільце Гольджі-Маццоні
Больові рецептори (ноцицептори) – вільні нервові закінчення. Шкірні нервові сплетення складаються з двох шарів і з верхнього шару до клітин епідермісу відходять у вигляді точок тонкі кінцеві волоконця. Гілки одного нервового волокна утворюють у шкірі мережу площею 1 см2. Мережі, що виникли при розгалуженні різних волокон, настільки тісно переплітаються один з одним, що сигнали дотику і болю йдуть відразу по кількох нервових шляхах. Подібні сплетення зустрічаються всюди - у шкірі, слизових оболонках, у внутрішніх органах. Найбільшу кількість ноцицепторів вдається виявити у шкірі та рогівці. У пахвових та пахвинних областях, а також у надниркових ямках число больових точок дорівнює 200 на 1 см2. На шкірі та на слизових оболонках можна знайти ділянки, що не сприймають біль при уколі, щипці, сильному тиску.

Больові рецептори та нервові волокна шкіри людини (схема)
У Останніми рокамивдалося виявити тонкі волоконця, що зв'язують вільні нервові закінчення з рецепторами дотику, тепла та холоду. Вони отримали назву волокон Тимофєєва. Наявністю цих волокон можна пояснити те що, що посилення тиску може викликати почуття болю. Для того щоб викликати почуття дотику в дотику, треба прикласти тиск 2-3 г на 1 мм2. А для того, щоб у тій же точці викликати біль, необхідний тиск у 200 г на 1 мм2.
Тільця Фатер-Пачіні (пластинчасті тільця) - капсульовані рецептори тиску (барорецептори) в округлій багатошаровій капсулі. Розташовуються в дермі, частіше - на межі дерми та гіподерми. Є швидкоадаптуються (реагують тільки в момент початку впливу), тобто реєструють силу тиску. Мають великі рецептивні поля, тобто представляють грубу чутливість. Розподілені в шкірі області пальців, зовнішніх статевих органів (крім того присутні в стінці сечового міхура, капсулі внутрішніх органів та ін.) Пластинчасті тільця - найбільші з усіх інкапсульованих нервових закінчень. Вони овальні, досягають 3-4 мм завдовжки і 2 мм завтовшки. Для них характерна наявність багатошарової пластинчастої сполучнотканинної оболонкою (зовнішньої колби), багатої гемокапілярами. Під сполучнотканинною оболонкою лежить зовнішня цибулина, що складається з 1060 концентричних пластинок, утворених сплощеними периневральними гексагональними епітеліоїдними клітинами. Увійшовши в тільце, нервове волокно втрачає мієлінову оболонку. Усередині тільця воно оточене лімфоцитами, які формують внутрішню цибулину.
Тельця Мейсснера – рецептори тиску (барорецептори), розташовані в дермі. Є шаруватою структурою з нервовим закінченням, що проходить між шарами. Є швидкоадаптуються. Мають малими рецептивними полями, тобто становлять тонку чутливість. Розташовані в сосочковому шарі шкіри пальців, губ, повік, статевих органів. Вони мають діаметр близько 100 мкм, зовні оточені сполучнотканинною капсулою. У складі цих тілець нейрогліальні клітини утворюють внутрішню колбу навколо кінцевого потовщення чутливого нервового волокна, яке розташовується паралельно поверхні шкіри.
Тільця Гольджі-Маццоні (цибулинні тільця) - рецептори тиску (барорецептори) - інкапсульовані чутливі нервові закінчення, що складаються з розгалуження чутливого нервового волокна, гліальної внутрішньої колби і сполучнотканинної капсули; зустрічається в шкірі, сполучній оболонці очного яблука, очеревині, кліторі, головці статевого члена, шкірі губ та країв рота, а також в інших покровах тіла.
Тельця (клітини) Меркеля – некапсульовані рецептори тиску (барорецептори). Є повільно адаптованими (реагують на всій тривалості впливу), тобто реєструють тривалість тиску. Мають малими рецептивними полями. Вони беруть участь у сприйнятті дотиків, оскільки тісно пов'язані з сіткоподібними кінцевими розгалуженнями чутливих нервів. Крім того, клітини Меркеля синтезують специфічні для нервових клітин маркери (нейрофіламенти, нейрональні адгезійні клітинні молекули та ін.). На підставі присутності в цитоплазмі клітин нейропептидів вони належать до дифузної ендокринної системи. Мет-енкефалін, що виробляється клітинами Меркеля, стимулює імунні реакції організму.
Рецептори волосяних цибулин - реагують на відхилення волосся.
Закінчення Руффіні - рецептори розтягування. Є повільноадаптованими, мають великі рецептивні поля. Реагують на зміщення шкіри, тепло (терморецептори) та тиск. Лежать у глибоких шарах шкіри, наприклад, підошви стопи. Діаметр - до 1 мм. Аферентне волокно утворює на кшталт кущика з немієлінізованих гілочок, які закінчуються колбоподібними терміналями (здуттями, оточеними леммоцитами). Закінчення щільно прилягають до фібробластів і колагенових волокон, що формують основу тільця. Сполучнотканинна капсула добре виражена.
Кінцеві колби Краузе – рецептори, що реагують на холод (терморецептори). Виявляються у кон'юнктиві, мові, зовнішніх статевих органах. Діаметр - до 150 мкм. Сферичні формою, мають тонку капсулу, численні розгалуження аферентного закінчення розташовуються у вигляді колби.
Загальна кількість температурних точок на поверхні шкіри дорослої людини дорівнює приблизно 280 тис., причому 30 тис. припадає на частку точок, що сприймають тепло. На поверхні тіла температурні точки розподілені дуже нерівномірно. Найбільш чутливі до температурних подразнень повіки очі, грудні залози, спина. Область чола мало чутлива до тепла і дуже сприйнятлива до холоду. Мало чутливі до різких теплових подразнень шкіра голови, нижніх кінцівок, слизова оболонка порожнини рота та язика.
Шкіра рясно забезпечена нервами і являє собою велике рецепторне поле, що сприймає подразнення, що надходять із зовнішнього та внутрішнього середовища.
Зважаючи на наявність багатого та різноманітного нервового апарату, шкіра відіграє важливу роль у життєдіяльності організму.
Нервовий апарат шкіри складається з нервових волокон та нервових закінчень, вільних або інкапсульованих. Вони розташовані переважно в дермі та епідермісі; значно менше їх у гіподермі.
Нервові стовбури, що проникають у шкіру, утворюють нервове сплетення в гіподермі. Від цього сплетення відходять нерви в дерму, утворюючи у ній нові сплетення. Від нервових сплетень гіподерми та дерми відходять нервові гілочки до волосяних мішечок, сальних та потових залоз, м'язів.
У шиловидному шарі епідермісу знаходяться спеціальні нервові апарати - клітини Меркеля, що сприймають тактильну чутливість, та вільні закінчення осьових циліндрів у вигляді загострень та гудзичних потовщень, що сприймають больову чутливість.

У сосочковому шарі дерми розташовані нервові закінчення, звані колби Краузе і тільця Мейсснера. Колби Краузе сприймають почуття холоду, їх багато в шкірі кистей, слизових оболонках, в ділянці клітора, головки статевого члена та внутрішнього листка крайньої плоті.

Тельця Мейсснера, що сприймають дотик, велику кількістьзнаходяться в шкірі долонної та бічної поверхонь пальців.

У шкірі налічується близько 500 000 клітин Меркеля та тілець Мейсснера.
У підшкірній жировій клітковині розташовані нервові прилади, що сприймають почуття тепла – тільця Руффіні та відчуття глибокого тиску – тільця Фатера – Пачіні.


На 1 см 2 шкіри припадає до 200 больових рецепторів, 20 тактильних 12 холодових та 2 теплових.
Слизова оболонка ротової порожнини вистелена багатошаровим плоским епітелієм, який значно відрізняється від епідермісу шкіри і майже не має рогового, блискучого і зернистого шарів. Епітелій фактично складається з базального та шилоподібного шарів.
Шилоподібний шар у верхній частині складається з кількох плоских, ніби здавлених клітин. Ороговіння епітелію в звичайних умовах не відбувається, за винятком ділянок на передній поверхні твердого піднебіння і верхівок ниткоподібних сосочків язика. Через відсутність трьох шарів епітелію дрібні численні кровоносні судини слизової оболонки ротової порожнини легко просвічуються і надають їй червоного кольору.
Строма (дерма) тут має таку ж будову, як і в шкірі, але в ній більше слинних, слизових, сальних, серозних (білкових) та змішаних залоз.
«Шкірні та венеричні хвороби»,
А.А.Студніцин, Б.Г.Стоянов
Рецепторні (чутливі) нервові закінчення сприймають сигнали із зовнішнього середовища (екстерорецептори) та внутрішніх органів (інтерорецептори). Залежно від природи подразнення, що реєструється рецепторами, вони поділяються відповідно до фізіологічної класифікації, на механорецептори, хеморецептори, терморецептори та больові рецептори (ноцицептори). У спеціалізованих органах почуттів (орган смаку, нюху, зору, рівноваги та слуху) є особливі рецепторні клітини, які сприймають відповідні подразнення.
Морфологічна класифікація чутливих нервових закінченьзаснована на особливостях їхньої структурної організації. Відповідно до цієї класифікації розрізняють вільні та невільні чутливі нервові закінчення; останні включають інкапсульовані та неінкапсульовані закінчення (рис. 8.18).
Мал. 8.18. Рецепторні (чутливі) нервові закінчення (по Rohen J.W., Lutjen-Drecoll E. 1982, із змінами). 1 - вільні нервові закінчення (СНТ) утворені термінальними розгалуженнями дендриту чутливого нейрона, що йде у складі нервового волокна (НВ). Невільні інкапсульовані нервові закінчення (2-4) утворені розгалуженнями дендриту, оточеними леммоцитами, разом із якими вони утворюють структуру, звану внутрішньої колбою (ВК). Зовні закінчення покриті сполучнотканинною капсулою (СТК). 2 - колба Краузе, 3 - дотик тільце (Мейснера), 4 - пластинчасте тільце (Фатер-Пачіні).
Вільні чутливі нервові закінченняскладаються тільки з термінальних розгалужень дендриту чутливого нейрона, вони зустрічаються в епітелії, а також у сполучній тканині. Проникаючи в епітеліальний пласт, нервові волокна втрачають мієлінову оболонку та нейролему, а базальна мембрана їх леммоцитів зливається з епітеліальною. Вільні нервові закінчення забезпечують сприйняття температурних (теплових та холодових), механічних та больових сигналів (рис. 8.19).

Мал. 8.19. Вільні нервові закінчення у сполучній тканині, представлені кінцевими розгалуженнями дендритів чутливих нейронів.
Невільні чутливі нервові закінчення містять усі компоненти нервового волокна. Вони поділяються на інкапсульовані (мають особливу сполучнотканинну капсулу) і неінкапсульовані.
Невільні неінкапсульовані нервові закінченняскладаються із розгалужень дендритів, оточених леммоцитами. Вони зустрічаються у сполучній тканині шкіри (дермі), а також власної платівки слизових оболонок.
Невільні інкапсульовані нервові закінченнядуже різноманітні, але мають єдиний загальний план будови: їх основу складають розгалуження дендриту, які безпосередньо оточені леммоцитами і зовні покриті особливою сполучнотканинною капсулою (див. рис. 8.18). До цього виду нервових закінчень відносять пластинчасті тільця (Фатер-Пачіні), дотик тільця (Мейснера) тільця Руффіні, колби Краузе, нервово-м'язові веретена і нервово-сухожильні веретена (сухожильні органи Гольджі).
Пластинчасті тільця (Фатер-Пачіні)зустрічаються у сполучній тканині внутрішніх органів та шкіри (рис. 8.20.). Вони мають вигляд округлих утворень діаметром 1-5мм, сприймають тиск та вібрацію. Структурними компонентами тільця є:
1) внутрішня колба (цибулина), утворена видозміненими сплощеними леммоцитами, в яку проникають одне або кілька нервових волокон, що мають прямий хід;
2) зовнішня колба - шарувата сполучнотканинна капсула, що складається з фібробластів і колагенових волокон, що утворюють 10-60 концентричних пластин, між якими є рідина.
При деформації пластин капсули тиск передається на нервове закінчення, що викликає деполяризацію мембрани.
 А А |  Б Б |
Мал. 8.20. На електронній мікрофотографії зображено: Тельця Фатер-Пачіні (РС) – інкапсульований рецептор. Капсула складається з паралельно розташованої платівки (мабуть, похідної Шванновської клітини), колагенових волокон. У центрі капсули розташовані розгалуження одного немієлінізованого нервового волокна (х100).
Дотик тільця (Мейснера)розташовані переважно у сосочковому шарі дерми, мають еліпсоїдну форму та невеликі розміри (близько 50-120 мкм) (рис. 8.21.). Їхня внутрішня колба складається з плоских гліальних клітин, що лежать перпендикулярно довгій осі тільця, між якими розташовуються гілочки дендритів. Між гліальними клітинами проникають колагенові фібрили, пов'язані з базальним шаром епітелію. Капсула тонка, перетворюється на периневрій.
 А А |  Б Б |
Мал. 8.21. Тельця Мейснера. Інкапсульований рецептор (М), що у шкірі губ, геніталій тощо. Має овальну форму, розташовується у шкірі прямо під епідермісом (Е). Рецептор складається з капсули, утвореної колагеновими волокнами, шванновськими клітинами (х320). (Б метод імпрегнації х150). У капсулі знаходяться кілька безм'якотних нервових закінчень, які рясно гілкуються.).
Тільця Руффінілежать у сполучнотканинній частині шкіри та капсулах суглобів; вони сприймають тиск і мають вигляд веретеноподібних структур завдовжки до 1-2 мм. Внутрішню колбу утворюють гліальні клітини, між якими розташовуються численні розгалужені терміналі дендритів з розширеннями на кінцях. Капсула добре виражена, утворена колагеновими волокнами.
Колби Краузе- дрібні (40-150 мкм) округлі тільця, які є механорецепторами та, можливо, холодовими рецепторами. Вони розташовані в сполучній тканині сосочкового шару дерми та власній платівці слизової оболонки порожнини рота, надгортанника, у кон'юнктиві ока. Внутрішня колба утворена сплощеними гліальними клітинами, між якими тонкі гілочки дендриту утворюють сплетення у вигляді клубочка. Капсула складається з плоских клітин, що є продовженням периневрії.
Нервово-м'язові веретени - рецептори розтягування волокон поперечносмугастих м'язів- складні інкапсульовані нервові закінчення, що мають як чутливу, так і рухову іннервацію. Число веретен у м'язі залежить від її функції і тим вище, чим більш точними рухами вона має. Нервово-м'язове веретено (рис. 8.22, 8.23.) має довжину 0,5-7 мм і розташовується паралельно ходу волокон м'яза, званих екстрафузальними (від лат. extra - поза і fuso - веретено, тобто розташованими за межами веретена) . Веретено покрите тонкою сполучнотканинною капсулою (продовженням периневрію), усередині якої знаходяться тонкі поперечно-смугасті інтрафузальні м'язові волокна двох видів:
Волокна з ядерною сумкою - у розширеній центральній частині яких містяться скупчення ядер (1-4 волокна/веретено);
Волокна з ядерним ланцюжком - більш тонкі з розташуванням ядер як ланцюжка у центральній частині (до 10 волокон/веретено).
Чутливі нервові волокна утворюють кільцеспіральні закінчення на центральній частині ішрафузальних волокон обох типів і гроноподібні закінчення біля країв волокон з ядерним ланцюжком.
Рухові нервові волокна - тонкі, утворюють дрібні нервово-м'язові синапси з обох боків інтрафузальних волокон, забезпечуючи їх тонус.

Мал. 8.22. Нервово-м'язове веретено. 1 - загальний вигляд веретена, що розташовується між екстрафузальними м'язовими волокнами (ЕФМЗ) та утвореного інтрафузальними м'язовими волокнами (ІФМВ), які оточені сполучнотканинною капсулою (СТК). 2 - деталі будови веретена, що містить два види ІФМВ: волокна з ядерною сумкою (ВЯС) та волокна з ядерним ланцюжком (ВЯЦ). Чутливі нервові волокна утворюють кільцеспіральні закінчення (КСВ) на центральній частині ІФМВ обох типів та гроноподібні закінчення (ГВО) біля країв ВЯЦ. Нервово-м'язове веретено містить також рухові нервові волокна та утворені ними нервово-м'язові синапси по краях ІФМВ (не показано).
 А А |  Б Б |
Мал. 8.23. Нервово-м'язове веретено N-нервове волокно, С – капсула. А – поздовжній зріз (х320), Б – поперечний.
Нервово-сухожильні веретени (сухожильні органи Гольджі) - рецептори розтягування- веретеноподібні інкапсульовані структури довжиною близько 0.5-1мм, що розташовуються в області з'єднання волокон поперечносмугастих м'язів з колагеновими волокнами сухожиль. Кожне веретено утворене капсулою з плоских фіброцитів (продовження периневрії), яка охоплює групу сухожильних пучків, обплетених численними термінальними гілочками нервових волокон, частково покритих леммоцитами. Порушення рецепторів виникає при розтягуванні сухожилля під час м'язового скорочення.
СПИСОК ЛІТЕРАТУРИ
ОСНОВНА ЛІТЕРАТУРА
1. Заварзін А.А. Основи порівняльної гістології. Л. ЛДУ, 1985. - 397 с.
2. Гістологія. За ред. Ю.І. Афанасьєва, Н.А.Юріной.- М.: Медицина, 1989.- 670 з.
3. Хем А., Кормак Д. Гістологія. (У 5 томах). М: Світ. 1982.
4. Антіпчук Ю.П. Гістологія із основами ембріології. - М:Освіта. 1983. - 240 с.
5. Антіпчук Ю.П. Гістологія з основами ембріології. - K.: Вища школа. 1976. - 141 с.
6. Волкова О.В., Єлецький О.М. Основи гістології із гістологічною технікою. -М.: Медицина, 1982. - 302 с.
7. Мануїлова Н.А. Гістологія із основами ембріології. М.: Просвітництво, 1973. - 214 с.
8. Троценко Б.В., Чирський Н.В. Навчальний посібник з курсу гістології (електронна версія). - Сімферополь, 2001.
9. Гістологія (введення в гістологію)/за ред. Е.Г. Улумбекова, Ю.А. Челещова. - М.: Геотар, 1997.
10. Гістологія/Ю.І. Афанасьєв, Н.А. Юрина, Б.В. Альошин. - М.: Медицина, 1989.
11. Гістологія/Ю.І. Афанасьєв, Н.А. Юрина, Є.Ф. Котовський та ін - М.: Медицина, 2002. -
12. Луцик О.Д., Іванова А.І. Кабак К.С. Гістологія людини. – Львів. Світ, 1992.
ДОДАТКОВА ЛІТЕРАТУРА
1. Єлісєєв В.Г. Гістологія М: Медицина, 1983.
2. Купріянов В.В. та ін. Судинний ендотелій. - К.: Здоров'я. - 248 с.
3. Абрамов М.Г. Гематологічний атлас. -М.: Медицина. 1985. - 237 с.
4. Шаде Дж., Форд Д. Основи неврології. М.: Мир, 1976. - 350 з.
5. Гістологія (Вступ до патології). За ред. Е.Г. Улумбекова, Ю.А. Челишева.-М.: Геогар. 1997. - 947 с.
6. Прохончук А.А. та ін Гомеостаз кісткової тканини в нормі та при екстремальному впливі. - М.: Наука, 1984. - 200 с.
7. Соколов В.Є. та ін Адаптивні властивості епітелію та його похідні. Атлас мікрофотографій. - М.: Наука, 1979. - 110 с.
8. Гурфінкель В.С., Левік Ю.С. Скелетний м'яз та функція. -М.: Наука, 1985. - 143 с.
9. Кауфман О.Я. Гіпертрофія та регенерація гладких м'язів. М.: Наука. 1979. - 183 с.
10. Клішов А.В. Гістогенез та регенерація тканин. - Л.: Медицина, 1984.-30 с.
11. Комісарчик Я.Ю., Миронов А.А. Електронна мікроскопія клітин та тканин. - Л.: Наука, 1990. - 140 с.
12. Заварзін А.А. Основи приватної цитології та порівняльної гістології багатоклітинних тварин. Л.: Наука, 1976.
13. Гацько Г.Г. Жирова тканина під час старіння. Мінськ. Наука та техніка, 1985. - 184 с.
14. Гаврилов О.К. Клітини кісткового мозку та периферичної крові -М.: Медицина, 1985. - 288 с.
15. Вельш У., Шторх Ф. Введення в цитологію та гістологію тварин. М: Мир.1976.
16. Виноградов В.В., Воробйова Н.Ф. Опасисті клітини, Новосибірськ, Наука, 1973.
17. Занябуш Л. Молекулярна та клітинна біологія. М: Світ. 1982. Т.3.
18. Зенгебуш П. Молекулярна біологія. М: Мир, т.1-3. 1982.
19. Саркісов Д.С. Нариси структурних основ гемостазу. М: Медицина, 1977.
20. Збірник «Молекули та клітини». За ред. Г.М.Франка. Випуски 1-5. М: Мир, 1966-1970.
21. Сєров В.В., Шехтер А.Б. Сполучна тканина. М: Медицина. 1981.
22. Касавіна Б.А., Торбенко В.П. Життя кісткової тканини. М: Наука. 1979.
23. Кассиль Г.М. Внутрішнє середовище організму. М: Наука. 1983. - 277 с.
24. Пігарэвський З.В. Зернисті лейкоцити та його властивості. М: Світ, 1978.
25. Пітерс А., Палей С., Вебстер С. Ультраструктура нервової системи. М: Світ. 1972.
26. Шубнікова Є.А. Лекції з гістології. М: МДУ, 1973.
27. Мозок (пер. з англ. під ред. Симонова П.В.) М: Мир. 1982.
28. Фріденштейн А.Я., Чортков І.М. Клітинні основи імунітету. М: Медицина. 1981.
Доповідь з біофізики:
"Механорецепція"
1. Призначення
Для активного існування в навколишньому середовищі, вищі організми мають велику кількість аналізаторів, що спеціалізуються на різних типахвпливу: на світі (зір), на звуку (слух), на смак і запах (нюх), на дотик і температуру (дотик), на гравітацію. Механорецепція входить до тями і разом з вестибулярним апаратам дозволяє найповніше уявити організму і свідомості положення кінцівок і тіла у просторі.
2. Функції
Аналізатор механорецепції, як і будь-який інший аналізатор, включає три просторово-функціональні частини: рецептор (отримання та трансформація механічного впливу в електричний імпульс), провідникові шляхи (передача імпульсу) та нервовий центр (аналіз отриманої інформації та формування еферентної відповіді). Усі ці частини забезпечують:
Сприйняття механічних впливів на шкіру: локалізація, напрямок руху, швидкість деформуючого джерела, його вібрації (тактильна рецепція);
- сприйняття механічних зрушень усередині органів та м'язів з метою визначення положення кінцівок та тіла у просторі (пропріоцепція);
- Сприйняття впливів на волосяний покрив.
- Трансформацію сигналу з механічного в електричний, який може бути передано нейронами.
- Передавання імпульсів з високою або низькою швидкістю до нервового центру.
- Формування у свідомості загальної картини положення тіла та кінцівок у просторі.
- Забезпечення вегетативної нервової системи інформації про положення тіла та його контроль (підтримання вертикального положення тіла при можливому засинанні в такому положенні, інформація про рухи очей уві сні).
3. Пристрій та принцип роботи.
Типові механорецептори, як правило, є інкапсульованими утвореннями. Деякі їх називають поверхневими кінцевими органами, оскільки вони у шкірі розташовані поверхнево. Це диски Меркеля, тільця Мейснера, тільця Фатера-Пачіні, тільця Догеля, колби Краузе, тільця Руффіні, нервово-сухожильні веретени, нервово-м'язові веретени та інші (Рис.1).
Рис.1. Різні види механорецепторів
Пластинчасті тільця (за старою термінологією тільця Фатера-Пачіні)розташовуються у сполучній тканині внутрішніх органів та глибоких шарах шкіри, особливо на подушечках пальців, на брижі, у молочній залозі, кишечнику та інших внутрішніх органах. Мають вигляд округлих утворень.
Структурними компонентами є:
Внутрішня колба (цибулина), утворена видозміненими леммоцитами, в яку проникають нервові волокна;
- зовнішня колба – шарувата сполучнотканинна капсула з фібропластів та колагенових волокон, що утворюють концентричні пластини, між якими є рідина.
Внутрішня частина капсули містить плоскі, концентрично розташовані нейрогліальні клітини, що обмежують внутрішню колбу. Рецепторне волокно входить у внутрішню колбу з одного його полюса та утворює контакти з гліальними клітинами. Термінальна частина рецепторного волокна містить дрібні сферичні мітохондрії та світлі синаптичні бульбашки. Зовнішній шар капсули складається з потужної сполучнотканинної оболонки, утвореної з плоских серповидних клітин і сполучнотканинних волокон, між якими знаходиться інтерстиціальна рідина. Пластинчасті тільця сприймають відчуття вібрації, натягу, тиску на органи та внутрішньоорганний тиск. Принцип роботи тілець Фатера-Пачіні на сьогодні погано вивчений. Сполучнотканинні пластинки та інтерстиціальна рідина капсули ймовірно сприяють посиленню тиску на нервове закінчення, внаслідок чого аксолемма деформується, змінюється її проникність та відбувається генерація потенціалу. Вважають, що місцем виникнення потенціалу дії в тільцях Пачіні є область першого перехоплення Ранв'є.
Дотик тільця (Мейснера)розміщуються в сосочковому шарі дерми, мають форму еліпсоїду і невеликі розміри. Це тактильні рецептори, що реагують на дотик. Присутня в дермі шкіри, особливо часто в кінчиках пальців, підошвах, сосках, повіках, губах та статевих органах. У центрі тільця Мейснера розташовується спірально згорнуте безмієлінове розгалуження мієлінового волокна, яке проходить через поперечно розташовані овальні клітини, що нагадують шванівські клітини. Зовні тільце покрите сполучнотканинною капсулою. Внутрішня колба складається з гліальних клітин, що лежать перпендикулярно довгій осі тільця, між якими розташовуються гілочки дендритів. Зовні знаходиться дуже тонка, переходить у периневрит шарувата капсула – зовнішня колба. Незначна деформація капсули передається гліоцитам і далі на дендрити.
Клітини Меркелярозташовані під епідермісом, мають великі неправильної форми ядри і мікроворсинки, що простягаються до епідермальних клітин. У їх підстав розміщуються дископодібні закінчення сенсорних аксонів (диски Меркеля). Група з 10 - 20 клітин Меркеля утворює синаптичні контакти із закінченнями одного сенсорного аксона (тільця Пінкуса-Ігго). Клітини Меркеля реагують на раптові усунення шкіри, наприклад, при погладжуванні.
Тільця Руффінілежать у сполучнотканинній частині шкіри та капсулах суглобів: вони сприймають тиск і мають вигляд веретеноподібних структур. Внутрішню колбу утворюють гліальні клітини, між якими розташовуються терміналі дендритів з розширеннями на кінцях. Капсула добре виражена.
Колби Краузе- дрібні округлі тільця, які є механорецепторами та холодовими рецепторами. Вони лежать у дермі шкіри, слизовій оболонці порожнини рота, підгортанника, у кон'юнктиві ока. Внутрішня колба утворена плоскими гліоцитами, між якими тонкі гілки дендриту утворюють сплетення у вигляді клубочка. Зовнішня капсула дуже тонка.
Генітальні тільця Догеля– перебувають у особливо чутливих областях шкіри (зовнішні статеві органи, молочні залози). Вони за будовою схожі на колби Краузе, але, на відміну від них, у тільце входить кілька відростків від нейроцитів, що викликає сильну іррадіацію збудження. Реагують на тиск імпульсами, що викликають статеве збудження.
Механочутливі вільні закінчення у шкірі. Є немієлінізованими аксонами, у них немає корпускулярних структур. До них відносяться рецептори волосяних фолікул, які реагують на тиск від руху волосся.
Нервово-м'язові веретени- рецептори розтягування поперечно-м'язів - нервові закінчення, що мають як чутливу, так і рухову іннервацію. Чутливі нервові волокна витками рясно обплітають кілька м'язових волокон, утворюючи навколо них подібність до муфти. У цій галузі м'язові волокна стоншуються, число міофібрил у них зменшується, а кількість ядер різко зростає. Нервово-м'язові веретени бувають оточені сполучнотканинною капсулою.
Рухові нервові волокна утворюють дрібні нервово-м'язові синапси по краях інтрафузальних волокон, забезпечуючи їх тонус, регулюючи довжину волокон. Всі вільний простірміж м'язовими волокнами заповнено рідиною та обмежено тонкою капсулою. Зміна тонусу м'яза веде до зміни тиску рідини та передається на дендрити. Кільцеспіральні закінчення реагують зміну довжини м'язового волокна і швидкість цієї зміни, гроздевидные закінчення – лише зміну довжини. Число веретен в м'язі залежить від її функції і тим вище, чим більш точними рухами вона має.
Нервово-сухожильні веретена – рецептори розтягування, розташовуються у місці з'єднання м'яза з сухожиллям, веретеноподібні структури завдовжки 0,5-1 мм. Кожне веретено має капсулу з фіброцитів, яка охоплює групу сухожильних пучків, обплетених закінченнями нервових волокон. Порушення рецепторів виникає при розтягуванні сухожилля під час м'язового скорочення.
Від рецептора імпульс рухається по рефлекторній дузі через спинний мозок і стовбур мозку до ядра таламуса і далі до кори.
4. Робочі характеристики
Кількість та щільність розміщення рецепторів:
Загальна кількість тактильних рецепторівоцінюється в 10 мільйонів штук, що згруповані в 1 мільйон аферентів.
- кількість вільних аферентних закінчень у шкірі: 50% від загальної кількості шкірних аферентів.
- Щільність розміщення вільних нервових закінчень у шкірі: 170 шт/см2
Розміри та інші структурні параметри:
Тельця Мейснера: 50-140 мкм
- Колби Крауза: 40-150 мкм
- Тільця Руффіні: 1-2 мм
- Тільця Фатера-Пачіні: 0.5-5 мм
- Нервово-сухожильні веретени:
- Розміри: 0.5-1 мм
- Кількість м'язових волокон у веретені: 15 штук
- Нервово-м'язові веретени
- Кількість м'язових волокон у веретені: 1-8 штук
- Діаметр виходять нервових волокон: 17 мкм (первинні) та 8 мкм (вторинні)
Швидкість передачі:
Психофізична оцінка ємності тактильних каналів: 5 біт/c
- Швидкість передачі імпульсів із вільних нервових закінчень: 1 м/c (С-волокна, IV тип)
- швидкість передачі імпульсів з інкапсульованих рецепторів: 50 м/c (А-волокна, II тип)
Роздільна здатність і пороги відчуттів:
Поріг відчуття шкірних рецепторів при натисканні: 10 мкм
- Одночасний просторовий поріг шкірних рецепторів:
- На губах та кінчиках пальців: 1-3 мм
- На спині, плечах, стегнах: 50-100 мм
- Послідовний просторовий поріг шкірних рецепторів:
- На губах та кінчиках пальців: 1 мм
- На спині, плечах, стегнах: 10-20 мм
- поріг відчуття вібрації: 150-300 Гц
- Мінімальна амплітуда вібрації, що відчувається: 1 мкм
Модальність(Специфічність до впливів):
Тільця Руффіні і колби Крауза крім механічних впливів детектують температуру, тобто. полімодальні. А тільця Мейснера можуть відчувати вібрацію. Також полімодальні неінкапсульовані вільні аференти.
- Інші рецептори унімодальні.
Рецепторні поля(Площа рецепції):
Для тактильних дисків площа дорівнює анатомічних розмірів.
- Для дисків Меркеля це агломерація із 30-50 дисків, що обслуговуються одним нервовим волокном.
- Для тілець Фатера-Паччіні ця область більша за анатомічний розмір.
Адаптація(Див. Таблиця 1).
Таблиця 1. Класифікація шкірних механорецепторів за швидкістю адаптації та адекватним стимулам.

5. Регуляція
Інформація від рецепторів передається до центральної нервової системи через спинальні або черепномозкові нерви. Відгалуження аксонів, що передають сенсорну інформацію, формують синаптичні закінчення на нейронах спинного мозку (або стовбура мозку у разі черепномозкових черв'яків), потім направляються у вищі відділи нервової системи (стволові ядра, які у свою чергу передають інформацію вище). У кожному з цих відділів нервової системи потік сенсорної інформації може фільтруватися - акцентуватися або, навпаки, блокуватися.
Сенсорні системи побудовані за ієрархічним принципом: сигнали від рецепторів надходять у нижчі рівніцентральної нервової системи (спинний мозок або стовбур мозку), звідки передаються до вищих відділів (ядра таламуса, кора великих півкуль, базальні ганглії). На кожній із цих послідовних стадій сенсорна інформація перетворюється, відбувається її фільтрація.
Потік інформації не односторонній, оскільки вищі відділи ієрархії посилають сигнали нижчі відділи. Крім того, сенсорна інформація обробляється не ланцюжком послідовних структур, а швидше багатьма областями мозку одночасно (або, як кажуть, паралельно). Паралельна обробка сенсорної інформації стає очевидною у вищих областях мозку, таких як кора півкуль. Тут окремі галузі спеціалізуються на обробці окремих елементів інформації.
Сенсорна інформація різноманітних не обробляється окремо. У багатьох областях мозку, званих асоціативними, відбувається змішання модальностей - наприклад, у парієтальній корі та пагорбах чотирихолмія. Нейрони у цих відділах мозку реагують різні стимули, наприклад, зорові, тактильні і слухові.
Структурами, що контролюють сенсорну інформацію, є кора великих півкуль (зокрема, префронтальна кора, яка відіграє важливу роль у контролі уваги), базальні ганглії, ретикулярна формація, таламус (зокрема ретикулярне ядро таламуса) та інші структури.
Одним із основних механізмів фільтрації сенсорної інформації є гальмування, яке виробляють ГАМК-ергічні синапси. Як правило, нейрон, що передає сенсорну інформацію, є збуджуючим. Фільтрують ці збуджуючі сигнали гальмівні нейрони. Гальмування може бути пресинаптичним (тобто блокуючим передачу сигналів за сенсорним аксоном до якогось нейрона) або постсинаптичним (гіперполяризуючий нейрон, що приймає сенсорні сигнали). Постсинаптичне гальмування дозволяє блокувати сигнали вибірково, так як клітина, що приймає, залишається здатною відповідати на інші, незаблоковані впливу.
На фільтрацію сенсорних сигналів впливають такі нейромедіатори як ацетилхолін, дофамін, ендорфіни та інші.
6. Енергозалежність
Оскільки весь аналізатор механорецепції є похідним нервової тканини, то енергозалежність є особливо високою. Нейрони постачаються поживними речовинамиі киснем через нейроглію та розташовані в ній кровоносні судини. При гіпоксії та тромбах може порушуватися харчування окремих ділянок (наприклад, шкіри або органів), почнеться ретроградна дегенерація нейронів (у деяких випадках можливе відновлення функцій волокна через деякий час після відновлення харчування) та втрату рецепції на ділянці. Однак гіпоксія в мозку може призвести до набагато гірших наслідків, втрати функцій цілих відділів або полів кори мозку.
7. Технічний аналог
Аналогом механорецепторів є пристрої, засновані на п'єзоефект. Тільця Руффіні у своєму складі мають колагенові волокна, а вони виявляють п'єзоелектричні властивості. П'єзоелектричний ефект (п'єзоефект) у тому, що з механічних деформації деяких кристалів у певних напрямах з їхньої гранях з'являються електричні заряди протилежних символів, тобто. механічні впливи трансформуються у напругу. Виходить, що тільця Руффіні функціонують як своєрідні п'єзоелектричні пристрої, оскільки в них між нервовою терміналлю та колагеновим волокном немає будь-якої іншої тканини. У техніці часто використовують зворотний п'єзоефект, коли електричний струм перетворюється на механічні коливання – наприклад, для генерації ультразвуку.
Список літератури
1. Шубнікова Є.А., Функціональна морфологія тканин. - М: Видавництво Московського університету. - 1981. - 326 с.
2. Фенькіна Р.П., Дегтярьов В.П., Коротич В.А., Навчальний посібник з нормальної фізіології. - М: МДМСУ. - 1994г
3. Сандаков Д.Б., Курс лекцій з фізіології. - Мінськ: біофак БДУ.
4. Шмідт Р., Основи сенсорної фізіології. - М: Світ. - 184. - 287 с.
5. Кузнєцов С.Л., Мушкамбаров Н.М., Горячкіна В.Л. Керівництво-атлас з гістології, цитології та ембріології.

Нічне або денне нетримання сечі – поширена, неприємна та дуже травмуюча проблема. Психіка дитини може...

В'язання гачком - один з найбільш цікавих видів рукоділля для майстрів по всьому світу. Різноманітність петель та...

Сумнівалася, чи ставити дані «твори», але на роботі колеги-мами зацікавилися, та й процес виготовлення я на...

Існує спеціалізований спортивний одяг, призначений для занять гірськолижним спортом.

Матеріали та інструменти: Пакувальний папір з білим візерунком та сітка; - широка шовкова стрічка білого кольору; - Тонка...

Яке ж новорічне свято без головних його героїв? Існують не лише тематичні вироби з паперу. Є навіть і...

Проводити лікування змовами від хвороб можуть лише люди, охрещені у православній церкві за правилами...

Придивляючись собі верхній одяг, звернула увагу на пухнасту коротко стрижену шубку. Її хутро було...

Сьогодні на полицях косметичних магазинів достаток – вибір засобів догляду, зокрема пудри для обличчя,...
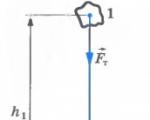
25.12.2014 Урок 32 (10 клас) Тема. Потенційна енергія 1. Робота сили тяжіння Обчислимо роботу,...

Опубліковано 01.09.17 08:41 Курбан Байрам у Москві 2017: святкова молитва проходить у Московській соборній...

Проте 29 серпня Володимир Путін, виступаючи із телезверненням до громадян країни, доручив пом'якшити збільшення...

Будь-який, навіть початківець з обробки металів знає, що, виконуючи токарні роботи на верстаті,...

Як навчитися красиво завивати волосся плойкою Пройшли ті часи, коли щоб накрутити волосся,...

В'язання гачком - один з найбільш цікавих видів рукоділля для майстрів по всьому світу. Різноманітність...

Сумнівалася, чи ставити дані «твори», але на роботі колеги-мами зацікавилися та й процес...